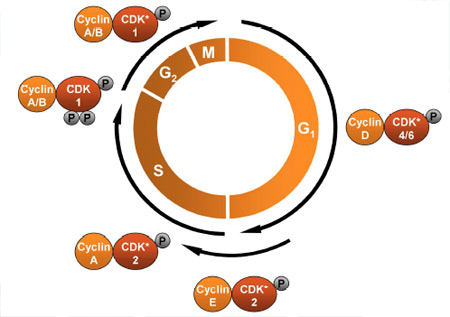
On two occasions I have been asked, "Pray, Mr. Babbage, if you put into the machine wrong figures, will the right answers come out?" ... I am not able rightly to apprehend the kind of confusion of ideas that could provoke such a question.
 Personally, I don't believe that enough experimental scientists appreciate the significance of sample extraction and preparation. After all, we place a great deal of trust in our dentist if an extraction is required! This is largely an experiential view and one that I was forced to confront when I first had to come to terms with transitioning basic to applied research in collaboration with a number of commercial organisations. The robustness of a method, such as ion exchange chromatography, electron microscopy, NMR spectroscopy etc., all stand or fall on the quality and quantity of the input sample. It doesn't matter how sophisticated your instrument is, if the sample under investigation does not meet a certain level of purity, or is present at too low a concentration, or has been prepared at the wrong pH, or salt concentration etc., you may as well not bother! As Arthur Kornberg himself commented: "Why waste pure thoughts on impure proteins!"
Personally, I don't believe that enough experimental scientists appreciate the significance of sample extraction and preparation. After all, we place a great deal of trust in our dentist if an extraction is required! This is largely an experiential view and one that I was forced to confront when I first had to come to terms with transitioning basic to applied research in collaboration with a number of commercial organisations. The robustness of a method, such as ion exchange chromatography, electron microscopy, NMR spectroscopy etc., all stand or fall on the quality and quantity of the input sample. It doesn't matter how sophisticated your instrument is, if the sample under investigation does not meet a certain level of purity, or is present at too low a concentration, or has been prepared at the wrong pH, or salt concentration etc., you may as well not bother! As Arthur Kornberg himself commented: "Why waste pure thoughts on impure proteins!"If part of your business is the sale of DNA purification kits, then you will not only sell on price, but on the simplicity, efficiency and reproducibility of your particular kit in producing samples fit for purpose in the downstream process. In most research labs, a method is often developed by an individual, which may subsequently becomes a key part of that particular laboratory's repertoire (often for several years). This process of method development and dissemination to the wider scientific community has been a pillar of experimental science in Universities and Research Institutes for many years (this is reflected in the many journals and books dedicated to experimental methodology). However, in our high-throughput, data-hungry world, the importance of personal experimental failure and improvement in the laboratory, has largely been sacrificed in the oncoming juggernaut which is our thirst for answers. Clearly, the current search for an effective Ebola vaccine, in the same way that Jonas Salk raced to deliver his polio vaccine, against a background of escalating human tragedy, focuses the mind in an important way. In many ways, the two competing forces: delivery of solutions and the provision of high quality scientific training are complementary: a good scientist knows when to slow down and take care at the bench, and when to buy an off the peg solution to move a project along. What I would advocate is that greater importance is placed on teaching robust sample preparation methods, as a means of mitigating some of the trouble-shooting shortcomings of many young scientists, in order that they are capable of making informed decisions in their work. The quality and quantity of the input sample in PCR (usually referred to as the "template" is critical for success.
 Moving on, amplification and magnification are generally associated with electronics and optics respectively. However, they both have the same general meaning. Consider a singer in a small room ( a typical classroom for example): the singer would normally be audible to everyone in that room. As the size of the room increases, for example a small assembly hall or a large theatre, the singer becomes inaudible, especially at the back of the room. Clearly this phenomenon limited the size of many concert venues, and at the same time stimulated architects to incorporate acoustic criteria into their designs. Nevertheless, there is always a point reached where the singer becomes inaudible to even someone with the keenest of ears! This not only stimulated architects, but also electronic engineers, and thus was born, the microphone: a device for capturing sound waves and converting them into an electrical current. Similarly an amplifier is an electronic device that increases the power of a signal, such as "amplifying" the normal level of sound from a solid body electric guitar. As a result of these two developments, it has become commonplace to hold concerts in football stadia, with suitable equipment. In an analogous way, it was impossible to observe single bacterial cells and certainly not viruses, until the development of optical and electron microscopes respectively. In the former, usually through a combination of optical lenses, the image of a small object can be obtained and otherwise "invisible" objects appear as "virtual" images. Electron microscopes use electrons as the source of illumination and can produce images around 5 000 times greater in magnification, owing to the difference in resolving power resulting in the use of electrons. PCR is similarly a method that does for DNA samples what the electron microscope does for imaging, or the modern microphone does for sound. So let's consider how PCR works.
Moving on, amplification and magnification are generally associated with electronics and optics respectively. However, they both have the same general meaning. Consider a singer in a small room ( a typical classroom for example): the singer would normally be audible to everyone in that room. As the size of the room increases, for example a small assembly hall or a large theatre, the singer becomes inaudible, especially at the back of the room. Clearly this phenomenon limited the size of many concert venues, and at the same time stimulated architects to incorporate acoustic criteria into their designs. Nevertheless, there is always a point reached where the singer becomes inaudible to even someone with the keenest of ears! This not only stimulated architects, but also electronic engineers, and thus was born, the microphone: a device for capturing sound waves and converting them into an electrical current. Similarly an amplifier is an electronic device that increases the power of a signal, such as "amplifying" the normal level of sound from a solid body electric guitar. As a result of these two developments, it has become commonplace to hold concerts in football stadia, with suitable equipment. In an analogous way, it was impossible to observe single bacterial cells and certainly not viruses, until the development of optical and electron microscopes respectively. In the former, usually through a combination of optical lenses, the image of a small object can be obtained and otherwise "invisible" objects appear as "virtual" images. Electron microscopes use electrons as the source of illumination and can produce images around 5 000 times greater in magnification, owing to the difference in resolving power resulting in the use of electrons. PCR is similarly a method that does for DNA samples what the electron microscope does for imaging, or the modern microphone does for sound. So let's consider how PCR works.

 By heating and cooling around 20-30 times, the amplification proceeds as 1 duplex becomes 2, 2 become 4, 4 become 8 etc. How many cycles are needed to achieve 1 000 000 fold amplification? This is made possible by the thermostability of the polymerase enzyme. Taq polymerase (LHS) was the first commercial enzyme used in PCR. Its technical suitability was accompanied by considerable financial success! The patents surrounding PCR (including the instrumentation) have not only been lucrative, but also highly controversial. Today, there are many polymerases to choose from, some are more accurate than others (higher fidelity), some are more robust, some are better for long amplicons etc. Depending on the application of PCR, there are many choices of kits and enzymes now available. We shall be working through the logistics of planning a PCR experiment prior to carrying out some amplifications in the lab after Easter.
By heating and cooling around 20-30 times, the amplification proceeds as 1 duplex becomes 2, 2 become 4, 4 become 8 etc. How many cycles are needed to achieve 1 000 000 fold amplification? This is made possible by the thermostability of the polymerase enzyme. Taq polymerase (LHS) was the first commercial enzyme used in PCR. Its technical suitability was accompanied by considerable financial success! The patents surrounding PCR (including the instrumentation) have not only been lucrative, but also highly controversial. Today, there are many polymerases to choose from, some are more accurate than others (higher fidelity), some are more robust, some are better for long amplicons etc. Depending on the application of PCR, there are many choices of kits and enzymes now available. We shall be working through the logistics of planning a PCR experiment prior to carrying out some amplifications in the lab after Easter.






.jpg)